|
| |||
|
Týdeník věnovaný aktualitám a novinkám z fyziky a astronomie. | |||
|
Udělení Nobelovy ceny za chemii pro rok 2015
Lucie Kulhánková
Když v roce 1953 James Watson s Francisem Crickem objasnili strukturu DNADNA – Deoxyribonucleic acid, deoxyribonukleová kyselina. Jde o nukleovou kyselinu, jejíž cukernou složkou je 2'-deoxyribóza. Báze jsou tvořeny především čtveřicí adenin, thymin, guanin a cytosin; vzájemně komplementární jsou nukleotidy s adeninem a thyminem, a nukleotidy s cytosinem a guaninem. DNA vytváří dvoušroubovici z navzájem komplementárních opačně orientovaných řetězců, pracovního a paměťového. Její hlavní funkcí je uchovávání genetické informace., nositelky genetického kódu, panoval všeobecný předpoklad, že musí jít o strukturu velice stabilní. Právě jeden z nositelů letošní Nobelovy cenyNobelova cena – je udílena švédskou Královskou akademií věd jednou ročně v pěti kategoriích: za fyziku, chemii, fyziologii a medicínu, literaturu a za úsilí o mír. Cena je hrazena z Nobelovy nadace, kterou založil Alfréd Nobel, vynálezce dynamitu, v roce 1895. První cena za fyziku byla udělena v roce 1901 Wilhelmu Roentgenovi za objev rentgenového záření. Hodnota Nobelovy ceny se mění, v roce 2021 činí 10 milionů švédských korun, tj. 25 milionů českých korun. Uděluje se vždy 10. prosince při výročí smrti Alfreda Nobela., Tomas Lindalh, demonstroval, že naše genetická informace je mnohem náchylnější k rozpadu a poškození, než jsme se domnívali. Každý den je poškozována ultrafialovým zářením, radiací, volnými radikály a dalšími procesy. Většina chyb v informaci, kterou DNA nese, zůstává bez projevu, nicméně i záměna jedné báze za jinou může způsobit závažné onemocnění případně smrt organizmu. Objev nestability DNA tak otevřel dveře k výzkumu procesů, jimiž se buněčný aparát snaží zabránit poškození. Právě za objevy molekulárních mechanizmů těchto oprav byla dne 7. října 2015 udělena Nobelova cena za chemii.

Molekula DNA. Zdroj: 100 000 Genomes Project.
|
DNA – Deoxyribonucleic acid, deoxyribonukleová kyselina. Jde o nukleovou kyselinu, jejíž cukernou složkou je 2'-deoxyribóza. Báze jsou tvořeny především čtveřicí adenin, thymin, guanin a cytosin; vzájemně komplementární jsou nukleotidy s adeninem a thyminem, a nukleotidy s cytosinem a guaninem. DNA vytváří dvoušroubovici z navzájem komplementárních opačně orientovaných řetězců, pracovního a paměťového. Její hlavní funkcí je uchovávání genetické informace. Enzymy – jednoduché či složené bílkoviny, které katalyzují chemické přeměny v živých organizmech. Endonukleázy – enzymy štěpící fosfodiesterovou vazbu uvnitř řetězce DNA. Některé štěpí DNA nespecificky. Často ale vyhledávají konkrétní sekvenci (nejčastěji krátký palindrom), těmto endonukleázám se pak říká restrikční endonukleázy. Mají široké využití v genovém inženýrství. Endonukleáza může tvořit konce DNA buď tupé, nebo lepivé, kde je DNA na obou vláknech štěpena nerovnoměrně. Exonukleázy – enzymy štěpící fosfodiesterovou vazbu na koncích řetězce nukleové kyseliny. Liší se podle rozeznávání 3' nebo 5' konce. Například 5'-3' exonukleáza štěpí od 5' konce a v buňkách se využívá při odstraňování 5' čepičky na mRNA. DNA polymeráza má také exonukleázovou aktivitu a je schopna štěpit v obou směrech, čímž umožňuje opravy DNA a proofreading. |
Fotoreaktivace
Fotoreaktivace byla prvním objasněným procesem opravujícím poškození DNADNA – Deoxyribonucleic acid, deoxyribonukleová kyselina. Jde o nukleovou kyselinu, jejíž cukernou složkou je 2'-deoxyribóza. Báze jsou tvořeny především čtveřicí adenin, thymin, guanin a cytosin; vzájemně komplementární jsou nukleotidy s adeninem a thyminem, a nukleotidy s cytosinem a guaninem. DNA vytváří dvoušroubovici z navzájem komplementárních opačně orientovaných řetězců, pracovního a paměťového. Její hlavní funkcí je uchovávání genetické informace.. Opravu buňka využívá na odstranění dimerůDimer – molekula složená ze dvou kovalentně vázaných podjednotek, takzvaných monomerů.. Ty vzniknou například při poškození UV zářením. Dimery pak nejsou schopny klasického párování, což často vede k potížím při replikaci. Klíčovým enzymemEnzymy – jednoduché či složené bílkoviny, které katalyzují chemické přeměny v živých organizmech. tohoto procesu je fotolyáza, která je schopná přeměnit energii absorbovaného fotonuFoton – základní kvantum energie elektromagnetického záření, polní částice elektromagnetické interakce. Má nulovou klidovou hmotnost a nemá elektrický náboj. Jeho energie a hybnost jsou přímo úměrné frekvenci záření (E = ħω, p = E/c). Stav fotonu zahrnuje také polarizaci, protože jde o příčné vlnění. Kvantování energie poprvé zavedl Max Planck při pokusech o vysvětlení záření černého tělesa. Albert Einstein dal těmto kvantům reálný význam v roce 1905 při vysvětlení fotoelektrického jevu. Samotný název foton poprvé pro tuto částici použil až americký fyzikální chemik Gilbert Lewis v dopise časopisu Nature z roku 1926. na energii chemickou a vytvořit volný radikál, s jehož pomocí rozštěpí thyminový dimer. K této reakci využívá fotolyáza anion redukovaného FADHFADH – flavinadenindinukleotid, prostetická čili kovalentně vázaná skupina mnoha proteinů. Existuje ve třech různých redoxních stavech, je tedy schopen přenášet elektrony, čehož se hojně využívá při Krebsově cyklu, kde vzniká jeho redukovaná forma, která se pak účastní i dýchacího řetězce.–, kofaktoruKofaktor – součást složených enzymů, neaminokyselinová skupina, která je nezbytně nutná pro funkci daného enzymu. Na ten se může vázat jak kovalentně, tak nekovalentně. Struktura kofaktorů je značně různorodá., který po excitaci fotonem přesune jeden ze svých elektronů na dimer, čímž vytvoří pár radikálů. Aniontový dimer se následně rozdělí a přebytečný elektron se vrátí zpět na nukleotid FADH•, který je v podobě aniontu FADH– připraven k dalším opravám. Celá oprava je proto závislá na světle. FADH– může přijmout energii i rezonančně od druhého chromoforuChromofor – světlocitlivá molekula, která absorbuje světelná kvanta s vhodnou vlnovou délkou. Tato kvanta nějakým způsobem změní stav molekuly – zkroutí ji, excitují, změní její vodivost atp. fotolyázy deazaflavinu, který funguje jako fotonová sběrná anténa.
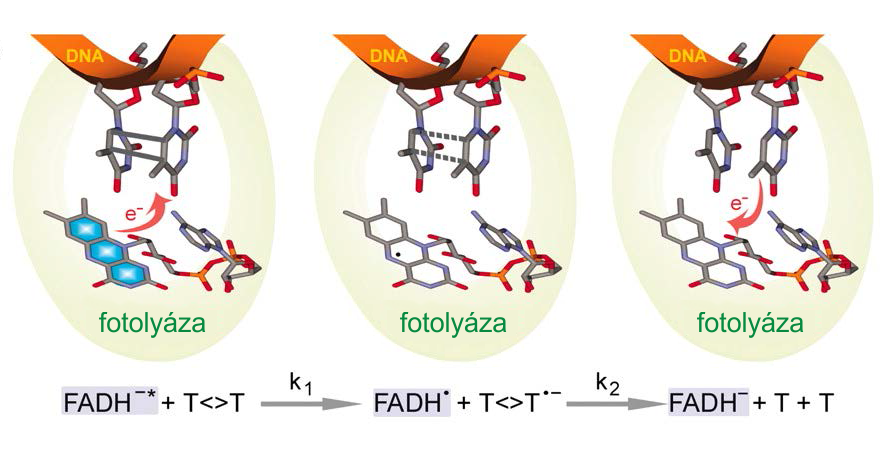
Zjednodušené schéma mechanizmu fotoreaktivace. Zdroj: Kao a kol., 2005.
Excizní opravy
Dimery samozřejmě nemohou zůstat ani v buňkách, které fotoreaktivaci nezvládají. Mezi takové buňky patří i buňky savců. Nicméně i zde jsou tyto cyklobutanové kruhy odstraňovány. Již v šedesátých letech si Richard Setlow a William Carrier všimli, že krátce po ozáření UV zářením zmizí dimeryDimer – molekula složená ze dvou kovalentně vázaných podjednotek, takzvaných monomerů. z genomickéGenom – veškerá genetická informace uložená v DNA (u některých virů v RNA) konkrétního organismu. Zahrnuje všechny geny a nekódující sekvence. DNADNA – Deoxyribonucleic acid, deoxyribonukleová kyselina. Jde o nukleovou kyselinu, jejíž cukernou složkou je 2'-deoxyribóza. Báze jsou tvořeny především čtveřicí adenin, thymin, guanin a cytosin; vzájemně komplementární jsou nukleotidy s adeninem a thyminem, a nukleotidy s cytosinem a guaninem. DNA vytváří dvoušroubovici z navzájem komplementárních opačně orientovaných řetězců, pracovního a paměťového. Její hlavní funkcí je uchovávání genetické informace. a přesunou se do frakcí DNA s menší molekulární hmotností. Tento poznatek naznačil, že dochází k vyštěpení celé části genetické informace a jejímu následnému dotvoření podle komplementárního vlákna. Molekulární mechanizmus tohoto procesu byl pak v roce 1983 objasněn Azizem Sancarem, jedním z letošních nositelů Nobelovy ceny.
Komplex dvou proteinů UvrA a jednoho UvrB skenuje DNA. Pokud UvrA nalezne distorzi v její struktuře, komplex se zastaví a UvrB pomocí své helikázovéHelikáza – enzym pohybující se podél fosfátové kostry a rozvíjející vlákna DNA od sebe. Proces je poháněn energií z ATP. Využívá se v mnoha klíčových buněčných procesech, jako jsou replikace, translace, rekombinace nebo opravy DNA. aktivity rozvine část dvoušroubovice dlouhou asi 5 párů bází. Následně UvrA od komplexu disociuje a zbývající UvrB podjednotka rekrutuje další enzymEnzymy – jednoduché či složené bílkoviny, které katalyzují chemické přeměny v živých organizmech. UvrC, tím se aktivuje endonukleázováEndonukleázy – enzymy štěpící fosfodiesterovou vazbu uvnitř řetězce DNA. Některé štěpí DNA nespecificky. Často ale vyhledávají konkrétní sekvenci (nejčastěji krátký palindrom), těmto endonukleázám se pak říká restrikční endonukleázy. Mají široké využití v genovém inženýrství. Endonukleáza může tvořit konce DNA buď tupé, nebo lepivé, kde je DNA na obou vláknech štěpena nerovnoměrně. aktivita UvrB, která naštěpí fosfátovou kostru DNADNA – Deoxyribonucleic acid, deoxyribonukleová kyselina. Jde o nukleovou kyselinu, jejíž cukernou složkou je 2'-deoxyribóza. Báze jsou tvořeny především čtveřicí adenin, thymin, guanin a cytosin; vzájemně komplementární jsou nukleotidy s adeninem a thyminem, a nukleotidy s cytosinem a guaninem. DNA vytváří dvoušroubovici z navzájem komplementárních opačně orientovaných řetězců, pracovního a paměťového. Její hlavní funkcí je uchovávání genetické informace. na 3' konci a UvrC štěpí na 5' konci (vysvětlení k číslování konců nalezne čtenář v AB 38/2009). Tak vznikne asi dvanáct bází dlouhý úsek, který je pomocí helikázyHelikáza – enzym pohybující se podél fosfátové kostry a rozvíjející vlákna DNA od sebe. Proces je poháněn energií z ATP. Využívá se v mnoha klíčových buněčných procesech, jako jsou replikace, translace, rekombinace nebo opravy DNA. UvrD odstraněn. Následně na DNA zůstává pouze UvrB, jehož vazbu zruší až vazba DNA polymerázyPolymerázy – enzymy, které podle jednoho vlákna DNA syntetizují k němu komplementární vlákno. Výchozí vlákno tedy pro tyto enzymy slouží jako jakási šablona, podle které je budováno nové vlákno. Svoji činnost však polymerázy nemohou zahájit v libovolném místě, potřebují, kousek již navázané komplementární nukleové kyseliny, primer, který začnou prodlužovat. Polymerázy prodlužují nové vlákno vždy ve směru od 5’ konce k 3’ konci nukleové kyseliny., která dosyntetizuje vyštěpené nukleotidy. Posledním krokem opravy je obnovení fosfátové kostry v místě štěpení, o které se postará enzym DNA ligázaDNA ligáza – enzym katalyzující vznik fosfodiesterové vazby mezi hydroxylovou skupinou 3' konce a fosfátovou skupinou 5' konce. ATP se váže v aktivním místě a štěpí se na AMP a pyrofosfát. Energie z této reakce je využita při tvorbě nové vazby. Využívá se například k ligování Okazakiho fragmentů při replikaci nebo při opravách jednovláknových i dvouvláknových zlomů..
Tvorba dimerů není jediným druhem poškození, který může v naší genetické informaci vzniknout. Poměrně běžnou záležitostí jsou také spontánní depurinaceDepurinace – odstranění purinu z organické molekuly. Purin je dusíkatá heterocyklická sloučenina. Je tvořena spojeným pyrimidinovým a imidazolovým kruhem. V DNA je zastoupen bázemi adeninem a guaninem. K purinům patří také kofein nebo kyselina močová. či depyrimidaceDepyrimidace – odstranění pyrimidinu z organické molekuly. Pyrimidin je dusíkatá heterocyklická sloučenina. Je tvořena pyrimidinovým kruhem. V DNA je zastoupen bázemi tyminem a cytosinem, v RNA se ještě navíc vyskytuje jako uracil., kdy DNADNA – Deoxyribonucleic acid, deoxyribonukleová kyselina. Jde o nukleovou kyselinu, jejíž cukernou složkou je 2'-deoxyribóza. Báze jsou tvořeny především čtveřicí adenin, thymin, guanin a cytosin; vzájemně komplementární jsou nukleotidy s adeninem a thyminem, a nukleotidy s cytosinem a guaninem. DNA vytváří dvoušroubovici z navzájem komplementárních opačně orientovaných řetězců, pracovního a paměťového. Její hlavní funkcí je uchovávání genetické informace. ztratí jednu bázi. Časté jsou také deaminaceDeaminace – odstranění aminoskupiny –NH2 z organické molekuly., alkylaceAlkylace – zavedení alkylové skupiny do organické molekuly. Alkyl je uhlovodíkový zbytek, nejčastěji se používá methyl (–CH3). a další poškození, která mohou při replikaci vyústit v chybné párováníKomplementární páry – vzájemně se doplňující dvojice částí molekul, které do sebe zapadají jako klíč do zámku nebo odlitek do formy. Umožňují samoorganizaci a vzájemné rozpoznávání na úrovni molekul. a zanesení mutace do další generace buněk. O opravu takovýchto poškození se starají enzymyEnzymy – jednoduché či složené bílkoviny, které katalyzují chemické přeměny v živých organizmech. glykosylázy. V našich buňkách existuje několik druhů těchto proteinůProteiny – bílkoviny, velké organické molekuly tvořené aminokyselinami uspořádanými do lineárních útvarů. Tyto dlouhé řetězce jsou pospojovány peptidovými vazbami (vazba karboxylové skupiny jedné molekuly a aminoskupiny druhé molekuly)., každý se pak specializuje na jiný druh oprav. Glykosylázy hledají nestandardní báze; pokud takový nukleotidNukleotidy – fosforylované nukleosidy, na pátý uhlík sacharidové složky je navázán jeden nebo více zbytků kyseliny fosforečné. Sacharidovou složku tvoří buď ribóza (ribonukleotidy) nebo deoxyribóza (deoxyribonukleotidy). Tvoří monomery nukleových kyselin, dále vstupují (přímo nebo ve formě derivátů) do enzymatických reakcí, přenosu energie a informace. Genetická informace je kódována posloupností nukleotidů. naleznou, vyklopí bázi do svého aktivního místa a štěpí glykosydickou vazbu mezi bází a deoxyribózouDeoxyribóza – přesněji 2’ deoxyribóza, derivát ribózy, ve které je hydroxylová skupina –OH na druhém uhlíku nahrazena vodíkem.. Enzym pak často zůstává vázán na DNA bez báze, dokud není nahrazen enzymem AP endonukleázou, který rozeznává apurinická a apyrimidická místa a štěpí fosfátovou kostruCukr-fosfátová kostra – základ nukleových kyselin, dlouhých polymerů skládajících se z neustále se opakujících podjednotek. Každá podjednotka se skládá z fosfátu, cukru (ribózy nebo deoxyribózy) a báze. Jednotlivé cukry jsou propojeny přes fosfáty fosfodiesterou vazbou vždy na 5' a 3' uhlíku. Touto vazbou je pak určena i směrovost DNA. na 5' konci, druhá vazba je pak štěpena fosfodiesterázou. Další funkcí AP endonukleázyEndonukleázy – enzymy štěpící fosfodiesterovou vazbu uvnitř řetězce DNA. Některé štěpí DNA nespecificky. Často ale vyhledávají konkrétní sekvenci (nejčastěji krátký palindrom), těmto endonukleázám se pak říká restrikční endonukleázy. Mají široké využití v genovém inženýrství. Endonukleáza může tvořit konce DNA buď tupé, nebo lepivé, kde je DNA na obou vláknech štěpena nerovnoměrně. je vazba na DNA polymerázuPolymerázy – enzymy, které podle jednoho vlákna DNA syntetizují k němu komplementární vlákno. Výchozí vlákno tedy pro tyto enzymy slouží jako jakási šablona, podle které je budováno nové vlákno. Svoji činnost však polymerázy nemohou zahájit v libovolném místě, potřebují, kousek již navázané komplementární nukleové kyseliny, primer, který začnou prodlužovat. Polymerázy prodlužují nové vlákno vždy ve směru od 5’ konce k 3’ konci nukleové kyseliny., která doplní chybějící bázi dle komplementárního vláknaKomplementární páry – vzájemně se doplňující dvojice částí molekul, které do sebe zapadají jako klíč do zámku nebo odlitek do formy. Umožňují samoorganizaci a vzájemné rozpoznávání na úrovni molekul.. Poté už opět stačí znovu vytvořit vazby na fosfátové kostře, o což se opětovně postará DNA ligázaDNA ligáza – enzym katalyzující vznik fosfodiesterové vazby mezi hydroxylovou skupinou 3' konce a fosfátovou skupinou 5' konce. ATP se váže v aktivním místě a štěpí se na AMP a pyrofosfát. Energie z této reakce je využita při tvorbě nové vazby. Využívá se například k ligování Okazakiho fragmentů při replikaci nebo při opravách jednovláknových i dvouvláknových zlomů..
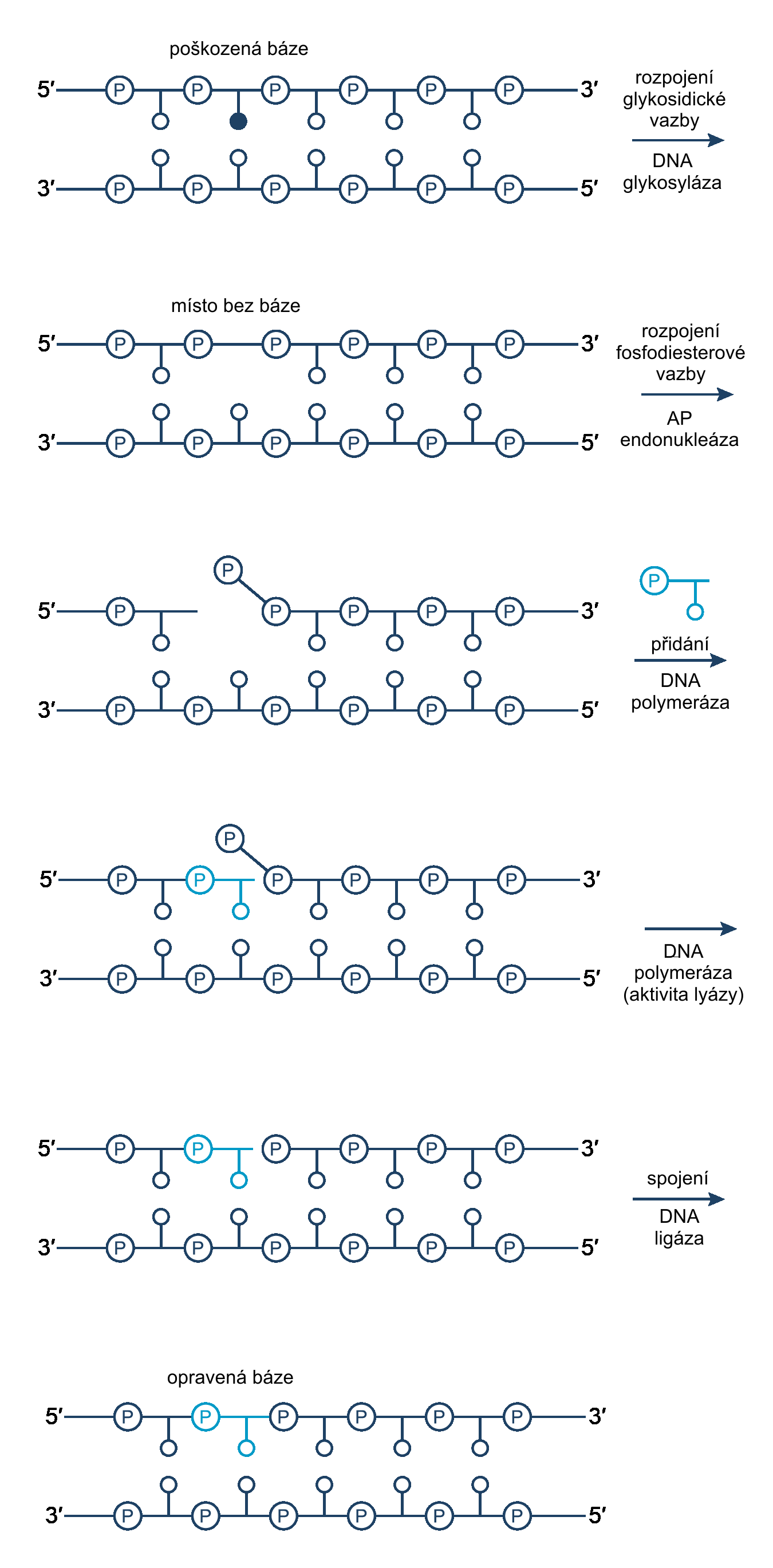
Schéma jednotlivých fází excizní opravy, při které je
odstraněn pouze jeden nukleotid. Zdroj: ATDBIO.
Opravy při replikaci
DNA polymerázaPolymerázy – enzymy, které podle jednoho vlákna DNA syntetizují k němu komplementární vlákno. Výchozí vlákno tedy pro tyto enzymy slouží jako jakási šablona, podle které je budováno nové vlákno. Svoji činnost však polymerázy nemohou zahájit v libovolném místě, potřebují, kousek již navázané komplementární nukleové kyseliny, primer, který začnou prodlužovat. Polymerázy prodlužují nové vlákno vždy ve směru od 5’ konce k 3’ konci nukleové kyseliny., enzymEnzymy – jednoduché či složené bílkoviny, které katalyzují chemické přeměny v živých organizmech., který replikuje DNADNA – Deoxyribonucleic acid, deoxyribonukleová kyselina. Jde o nukleovou kyselinu, jejíž cukernou složkou je 2'-deoxyribóza. Báze jsou tvořeny především čtveřicí adenin, thymin, guanin a cytosin; vzájemně komplementární jsou nukleotidy s adeninem a thyminem, a nukleotidy s cytosinem a guaninem. DNA vytváří dvoušroubovici z navzájem komplementárních opačně orientovaných řetězců, pracovního a paměťového. Její hlavní funkcí je uchovávání genetické informace. při dělení buněk, má sama o sobě exonukleázovouExonukleázy – enzymy štěpící fosfodiesterovou vazbu na koncích řetězce nukleové kyseliny. Liší se podle rozeznávání 3' nebo 5' konce. Například 5'-3' exonukleáza štěpí od 5' konce a v buňkách se využívá při odstraňování 5' čepičky na mRNA. DNA polymeráza má také exonukleázovou aktivitu a je schopna štěpit v obou směrech, čímž umožňuje opravy DNA a proofreading. aktivitu, která jí umožňuje případný chybně zařazený nukleotidNukleotidy – fosforylované nukleosidy, na pátý uhlík sacharidové složky je navázán jeden nebo více zbytků kyseliny fosforečné. Sacharidovou složku tvoří buď ribóza (ribonukleotidy) nebo deoxyribóza (deoxyribonukleotidy). Tvoří monomery nukleových kyselin, dále vstupují (přímo nebo ve formě derivátů) do enzymatických reakcí, přenosu energie a informace. Genetická informace je kódována posloupností nukleotidů. vyštěpit a vzniklou chybu opravit. Při pouze tomto druhu oprav by pravděpodobnost chyby na jeden zápis (frekvence chyb) byla asi 5×10−5. Nicméně odhadovaná frekvence chyb v našem genomuGenom – veškerá genetická informace uložená v DNA (u některých virů v RNA) konkrétního organismu. Zahrnuje všechny geny a nekódující sekvence. je přibližně 1×10−8. Od tohoto poznatku pak zbýval už jen krok k objevu takzvané mismatch repair. K objasnění tohoto procesu velkým dílem přispěl poslední nositel Nobelovy ceny, Paul Modrich. Jedním z problémů, který musí opravný mechanizmus vyřešit, je to, které vlákno má opravit, který z nesprávně párujících nukleotidů je zařazen špatně. V tomto případě se starý řetězec od nového pozná podle metylacíMetylace – připojení metylového zbytku -CH3 k aminokyselině v proteinu., které se běžně vyskytují na obou vláknech DNA v konkrétních sekvencích (například v Escherichia coliEscherichia coli – gramnegativní baktérie tyčinkovitého tvaru, běžně osidlující tlusté střevo teplokrevných živočichů, včetně člověka. Je fakultativně anaerobní, což znamená, že v případě potřeby dokáže převést svůj metabolismus přizpůsobený na přítomnost kyslíku na fermentaci za nepřítomnosti kyslíku. Je významná z hygienického hlediska jako jeden z indikátorů fekálního znečištění pitné vody. Její přítomnost ve střevě je pro organismus obvykle prospěšná, může však nést genetickou výbavu, která z ní činí původce závažných střevních onemocnění. Vzhledem ke svým nenáročným růstovým vlastnostem a dobře prozkoumané DNA definovaných kmenů je oblíbeným modelovým organismem či nosičem cizorodých genů. jde o sekvenci bází GATC), ovšem nové vlákno ještě nestihlo být metylováno. Nicméně tento systém funguje v Escherichia coli, ale způsob rozeznání správného vlákna v eukaryotických buňkáchEukaryota – jedna ze tří domén (nadříší) organizmů (archea, bacteria, eukaryota), jednobuněčné i vícebuněčné organizmy, které obvykle mají obsah buňky rozdělený membránami na oddělené prostory a genetickou informaci umístěnou v podobě lineárních řetězců DNA v formovaném jádře. Rozmnožují se pohlavně i nepohlavně. je stále neznámý. V Escherichia coli byl objasněn molekulární mechanizmus mismatch repair, kde proteinProteiny – bílkoviny, velké organické molekuly tvořené aminokyselinami uspořádanými do lineárních útvarů. Tyto dlouhé řetězce jsou pospojovány peptidovými vazbami (vazba karboxylové skupiny jedné molekuly a aminoskupiny druhé molekuly). MutS rozeznává páry, které nepárují dle Watson-Crickova párováníKomplementární báze – doplněk dané báze do Watsonova-Crickova páru. Pro A je to v DNA T, v RNA U, pro C je komplementární G, pro G naopak C, pro T, případně U, je to A.. Po rozeznání další enzym MutH váže nejbližší nemetylované sekvence GATC, čímž určí orientaci dalších kroků. Protein MutL, který funguje jako mediátor mezi MutS a MutH, předá signál od MutS, a tím aktivuje endonukleázovouEndonukleázy – enzymy štěpící fosfodiesterovou vazbu uvnitř řetězce DNA. Některé štěpí DNA nespecificky. Často ale vyhledávají konkrétní sekvenci (nejčastěji krátký palindrom), těmto endonukleázám se pak říká restrikční endonukleázy. Mají široké využití v genovém inženýrství. Endonukleáza může tvořit konce DNA buď tupé, nebo lepivé, kde je DNA na obou vláknech štěpena nerovnoměrně. aktivitu MutH, která naštěpí nascentní vlákno poblíž nemetylované oblasti. Následně dojde k interakci s helikázouHelikáza – enzym pohybující se podél fosfátové kostry a rozvíjející vlákna DNA od sebe. Proces je poháněn energií z ATP. Využívá se v mnoha klíčových buněčných procesech, jako jsou replikace, translace, rekombinace nebo opravy DNA. UvrD, která separuje vlákna DNA od sebe. Vytlačování vlákna pak pokračuje a zastaví se až za chybou. Následně je pak vlákno znovu dosyntetizováno pomocí polymerázyPolymerázy – enzymy, které podle jednoho vlákna DNA syntetizují k němu komplementární vlákno. Výchozí vlákno tedy pro tyto enzymy slouží jako jakási šablona, podle které je budováno nové vlákno. Svoji činnost však polymerázy nemohou zahájit v libovolném místě, potřebují, kousek již navázané komplementární nukleové kyseliny, primer, který začnou prodlužovat. Polymerázy prodlužují nové vlákno vždy ve směru od 5’ konce k 3’ konci nukleové kyseliny..
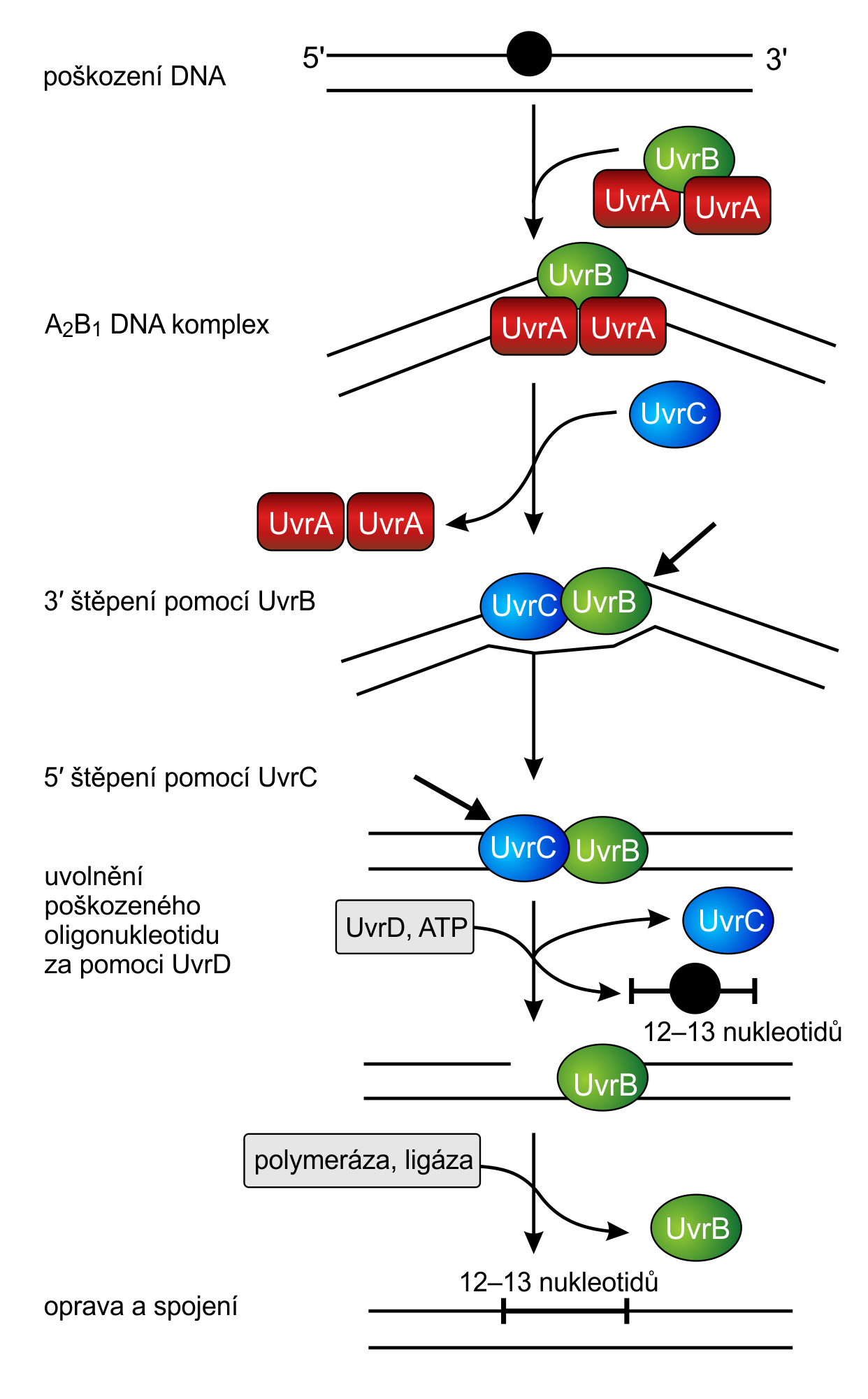
Princip excizní opravy, kde je vystřižen celý úsek DNA. Jde o
takzvanou opravu NER
(Nucleotide Excision Repair). Zdroj: [2].
Tomas Lindahl (1938)
Tomas Lindahl se narodil 28. ledna 1938 ve švédském Stockholmu. V roce 1967 získal doktorát na Institutu Karolinska ve Stockholmu, o tři roky později pak na stejném místě získal i lékařský titul. Ve studiích pokračoval v Princetonu a na Rockefellerově univerzitě ve Spojených státech. V letech 1978 až 1982 působil jako profesor lékařské chemie na Gothenburgské univerzitě. V roce 1981 se přestěhoval do Velké Británie. Mezi lety 1986 a 2005 byl ředitelem laboratoří Clare Hall, kde až do roku 2009 pokračoval ve svém výzkumu oprav DNA. V současné době je emeritním vedoucím skupiny. V roce 2007 byl oceněn Královskou medailí za fundamentální přínos k porozumění oprav DNA. V roce 2010 získal Copleyho medaili. V roce 2015 obdržel nejvyšší možné ocenění – Nobelovu cenu za výzkum mechanických oprav DNA. O tuto cenu se dělí s Paulem Modrichem a Azizem Sancarem.

Tomas Lindahl (1938)
Paul Modrich (1946)
Paul Modrich je americký biochemik narozený dne 13. června roku 1946. Narodil se v malém městečku Raton v Novém Mexiku. Zajímavostí může být, že jeho prarodiče emigrovali do Spojených států z Chorvatska. Vystudoval biologii na Massachusettském technologickém institutu, kde promoval roku 1968. Doktorát získal o pět let později za biochemii na Stanfordu. Profesorem se stal na Duke University, kde působí dodnes. Je členem Americké akademie umění a vědy. V roce 2015 obdržel nejvyšší možné ocenění – Nobelovu cenu za výzkum mechanických oprav DNA. O tuto cenu se dělí s Tomasem Lindahlem a Azizem Sancarem.

Paul Modrich (1946)
Aziz Sancar (1946)
Aziz Sancar je turecký biochemik působí ve Spojených státech. Narodil se dne 8. září 1946 v Savuru na jihovýchodě Turecka. Jeho rodiče neuměli číst, ale podporovali Sancara v získání vzdělání. V roce 1969 dokončil studium medicíny v Istanbulu, doktorát získal v Americe na Texaské univerzitě v Dallasu. Za svůj život obdržel mnoho ocenění, například medaili Národního institutu pro zdraví či tureckou Koç cenu. V současné době je profesorem biochemie a biofyziky na Univerzitě Severní Karolíny. V roce 2015 obdržel nejvyšší možné ocenění – Nobelovu cenu za výzkum mechanických oprav DNA. O tuto cenu se dělí s Tomasem Lindahlem a Paulem Modrichem.

Aziz Sancar (1946)
Odkazy
- Nobel Prize homepage: The Nobel Prize in Chemistry 2015 – The cells’ toolbox for DNA repair, 7 Oct 2015
- The Royal Swedish Academy of Sciences: Scientific Background on the Nobel Prize in Chemistry 2015 – Mechanistic Studies of DNA Repair, compiled by the Class for Chemistry of the Royal Swedish Academy of Sciences; 7 Oct 2015
- Atdbio: Mutagenesis and DNA repair
- Vítězslav Kříha: Nanotruhličky z DNA; AB 38/2009
- Vítězslav Kříha: Nanoburlaci – kráčející molekuly I: Inspirace v buňce; AB 7/2010
- Vítězslav Kříha: Nanoburlaci – kráčející molekuly II: DNA nanorobot, AB 15/2010
![]()
![]()
