|
| |||
|
Týdeník věnovaný aktualitám a novinkám z fyziky a astronomie. | |||
|
Nanotruhličky z DNA
Vítězslav Kříha
Není nad to mít pořádnou truhlu či truhličku. Vhodně prostornou, se solidními stěnami, víkem a především zámkem, který ji umožní otevřít na tom správném místě ve správnou chvíli. Jakmile máme truhlu, můžeme do ní schovávat poklady, můžeme si do ní sbalit věci na cestu či na později, a k uložení nebezpečného obsahu je truhla také k nezaplacení. V nanosvětě tomu není jinak. Co s neposednými molekulami, když nenastal jejich čas a nejsou na svém místě? Zavřít je do truhly 42×36×36 nm3, vytvořené z DNADNA – Deoxyribonucleic acid, deoxyribonukleová kyselina. Jde o nukleovou kyselinu, jejíž cukernou složkou je 2'-deoxyribóza. Báze jsou tvořeny především čtveřicí adenin, thymin, guanin a cytosin; vzájemně komplementární jsou nukleotidy s adeninem a thyminem, a nukleotidy s cytosinem a guaninem. DNA vytváří dvoušroubovici z navzájem komplementárních opačně orientovaných řetězců, pracovního a paměťového. Její hlavní funkcí je uchovávání genetické informace., a nechat je pod zámkem, dokud nebudou otevřeny nanoklíčem, kterým je opět kousek vhodné DNA.
|
Heterocyklické sloučeniny – organické sloučeniny vytvářející cyklické struktury, ve kterých jsou do kruhu vázány nejen atomy uhlíku, ale i jiné atomy, nejčastěji kyslík, dusík nebo síra. Nejstabilnější a nejvýznamnější jsou pěti a šestičlenné heterocyklické sloučeniny. Cyklus v molekule může být jeden, časté jsou i sloučeniny s více cykly, které většinou sdílejí část atomů v sousedících cyklech (kondenzované cykly). Nukleové báze – báze nukleových kyselin, heterocyklické struktury nesoucí informaci v nukleových kyselinách. Vedle nukleových kyselin jsou součástí sloučenin hrajících důležitou roli v enzymatických reakcích a přenosu energie a informace v buňce. Dělí se na purinové a pyrimidinové báze. Vedle pěti majoritních bázi adeninu (A), guaninu (G), thyminu (T), cytosinu (C) a uracilu (U) existuje řada minoritních bází. Monosacharidy – organické sloučeniny tvořené kostrou vzájemně vázaných uhlíků, kde k jednomu uhlíku je vázán kyslík dvojnou vazbou (karbonylová skupina C=O) a k ostatním je vždy po jednom navázán kyslík v podobě hydroxylové skupiny –OH. Pokud je uhlík vázaný s kyslíkem dvojnou vazbou na konci řetězce, nazývají se monosacharidy aldózy, pokud je uvnitř řetězce, hovoříme o ketózách. Podle počtu uhlíků dělíme monosacharidy na triózy (3 C), tetrózy (4 C), pentózy (5 C), hexózy (6 C) a heptózy (7 C). Karbonylová skupina C=O je vysoce reaktivní, což u delších řetězců, pentóz a zejména hexóz, umožňuje vytváření heterocyklické formy monosacharidů díky reakci s –OH skupinami navázanými na vzdálenějších uhlících. Tato cyklická forma je v rovnováze s formou, ve které se vyskytuje karbonylová skupina. Anomerní uhlík monosacharidů – hemiacetalový nebo hemiketalový uhlík monosacharidů, uhlík, který v monosacharidu vstupuje do karbonylové skupiny C=O, která může vytvářet cyklickou formu sacharidu. Glykosidy – deriváty monosacharidů, ve kterých je na anomerní uhlík navázán náhradou za hydroxylovou skupinu jiný složitější substituent. V případě, kdy je substituent navázán přes kyslík, hovoříme O-glykosidech, pokud přes dusík, N-glykosidech (glukosaminech) a pokud přes síru, S-glykosidech. O-glykosidovou vazbou jsou monosacharidy vzájemně vázány do oligosacharidů a polysacharidů. Ribóza – monosacharid, aldóza tvořená pěti uhlíky. Deoxyribóza – přesněji 2’ deoxyribóza, derivát ribózy, ve které je hydroxylová skupina –OH na druhém uhlíku nahrazena vodíkem. |
|
Nukleosidy – glykosidy ribózy (ribonukleosidy) nebo deoxyribózy (deoxyribonukleotidy), na něž jsou N-glykosidovou vazbou navázány nukleové báze dusíkem v heterocyklu. Nukleotidy – fosforylované nukleosidy, na pátý uhlík sacharidové složky je navázán jeden nebo více zbytků kyseliny fosforečné. Sacharidovou složku tvoří buď ribóza (ribonukleotidy) nebo deoxyribóza (deoxyribonukleotidy). Tvoří monomery nukleových kyselin, dále vstupují (přímo nebo ve formě derivátů) do enzymatických reakcí, přenosu energie a informace. Genetická informace je kódována posloupností nukleotidů. Nukleové kyseliny – makromolekuly zajišťující v živé buňce ukládaní, přenos a zpracování genetické informace, jsou lineární orientované polymery tvořené nukleotidy. Informační obsah nukleotidů nesou heterocyklické dusíkaté báze odvozené buď od purinu, nebo od pyrimidinu. V ribonukleotidech, vytvářejících ribonukleovou kyselinu (RNA) jsou purinové báze adenin (A) a guanin (G) a pyrimidinové báze cytosin (C) a uracil (U) navázány v místě uhlíku 1’ na ribóza 5-fostát. V deoxyribonukleotidech, vytvářejících deoxyribonukleovou kyselinu (DNA) je na uhlíku 2’ nahrazena hydroxylová skupina vodíkem. DNA využívá purinové báze A a G, avšak pyrimidinové báze cytosin (C) a thymin (T). Komplementární páry – vzájemně se doplňující dvojice částí molekul, které do sebe zapadají jako klíč do zámku nebo odlitek do formy. Umožňují samoorganizaci a vzájemné rozpoznávání na úrovni molekul. Watsonovy-Crickovy páry – W-C páry, komplementární páry nukleotidů v nukleových kyselinách. Komplementaritu v párech nukleotidů zajišťují báze nukleových kyselin. V DNA jsou komplementárními páry (A,T) a (C,G), pro RNA (A,U) a (C,G). Komplementární báze – doplněk dané báze do Watsonova-Crickova páru. Pro A je to v DNA T, v RNA U, pro C je komplementární G, pro G naopak C, pro T, případně U, je to A. Polymerázy – enzymy, které podle jednoho vlákna DNA syntetizují k němu komplementární vlákno. Výchozí vlákno tedy pro tyto enzymy slouží jako jakási šablona, podle které je budováno nové vlákno. Svoji činnost však polymerázy nemohou zahájit v libovolném místě, potřebují, kousek již navázané komplementární nukleové kyseliny, primer, který začnou prodlužovat. Polymerázy prodlužují nové vlákno vždy ve směru od 5’ konce k 3’ konci nukleové kyseliny. PCR – Polymerase Chain Reaction, polymerázová řetězová reakce, metoda zmožení části vlákna DNA mezi dvěma známými sekvencemi nukleotidů opakovaným rozpojováním každé dvouvláknové DNA na dvě jednovláknové DNA a dotvářením podle nich nových komplementárních řetězců do dvouvláknové DNA. DNA počítače – řešení paralelní úloh pomocí párování komplentárních úseků DNA. DNA origami – samoorganizující se 2D a 3D objekty, které díky přilnavosti většího množství vzájemně komplementárních úseků DNA zaujmou požadovaný tvar a vytvářejí příčné můstky mezi dvoušroubovicemi DNA. |
Stavební kameny, struktura DNA, kopírování definovaných úseků DNA
Deoxyribonukleová kyselina v přírodě vytváří buď dlouhé lineární molekuly tvořené dvoušroubovicí dvou vláken, nebo se konce této dvoušroubovice uzavírají samy do sebe a vytváří prstenec. Výchozími stavebními kameny DNA jsou purinové nukleové báze adenin a guanin a pyrimidinové nukleové báze cytosin a thymin, navázané N-glykosidovou vazbou na deoxyribózu.
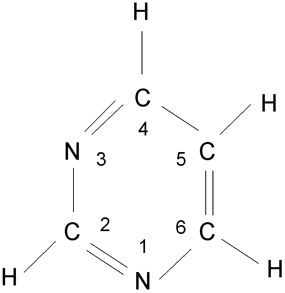
Heterocyklická sloučenina pyrimidin, číslování atomů v pyrimidinových bázích C, U, T.
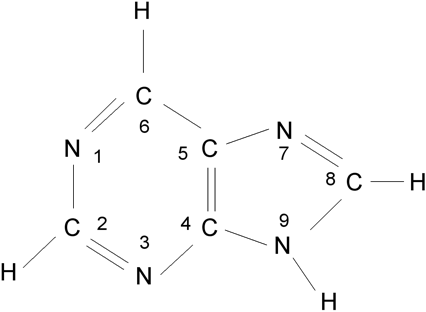
Purin, dva kondenzované heterocykly, číslování atomů v purinových bázích A, G.
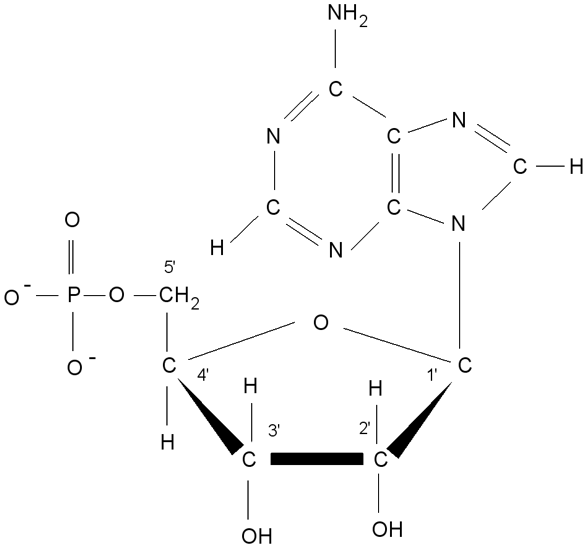
Ribonukleotid adenosinmonofosfát, číslování v molekule ribózy
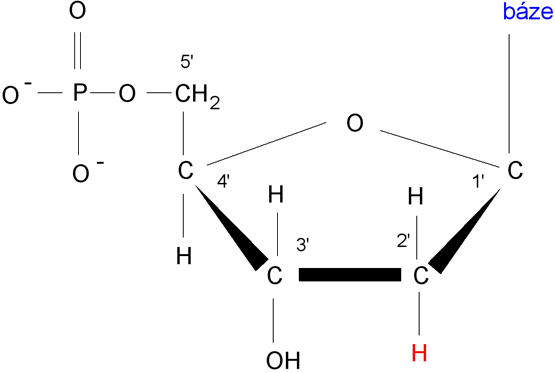
Deoxyribonukleotid s obecnou bází, číslování uhlíků v molekule
deoxyribózy,
povšimněte si změny na uhlíku 2’.
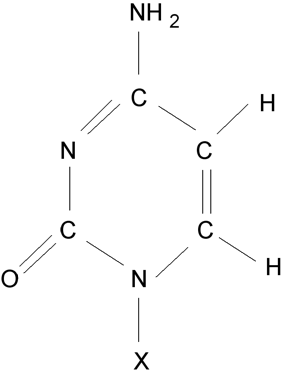
Cytosin (X odpovídá vodík), cytidinnukledotid (v místě X je vázán uhlík 1’ cyklu ribózy/deoxyribózy).
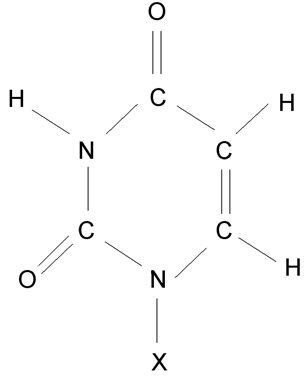
Uracil (X odpovídá vodík), uridinnukledotid (v místě X je vázán uhlík 1’ cyklu ribózy).
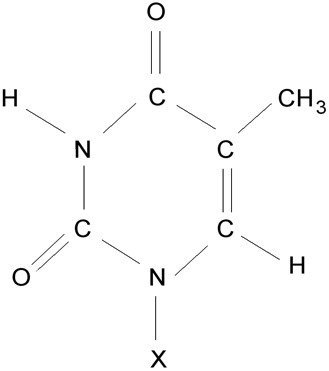
Thymin (X odpovídá vodík), thymidinnukledotid (v místě X je vázán uhlík 1’ cyklu deoxyribózy).
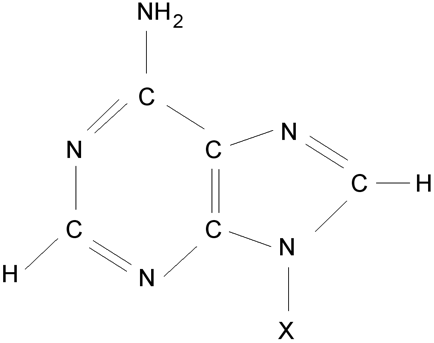
Adenin (X odpovídá vodík), adenosinnukledotid
(v místě X je vázán uhlík 1’ cyklu ribózy/deoxyribózy).
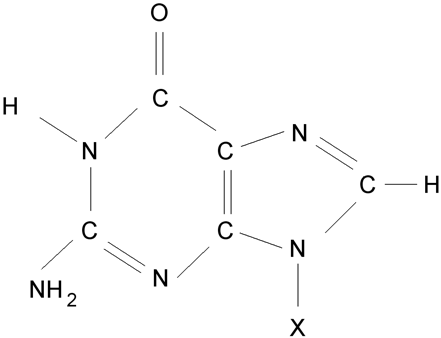
Guanin (X odpovídá vodík), guanosinnukledotid
(v místě X je vázán uhlík 1’ cyklu ribózy/deoxyribózy).
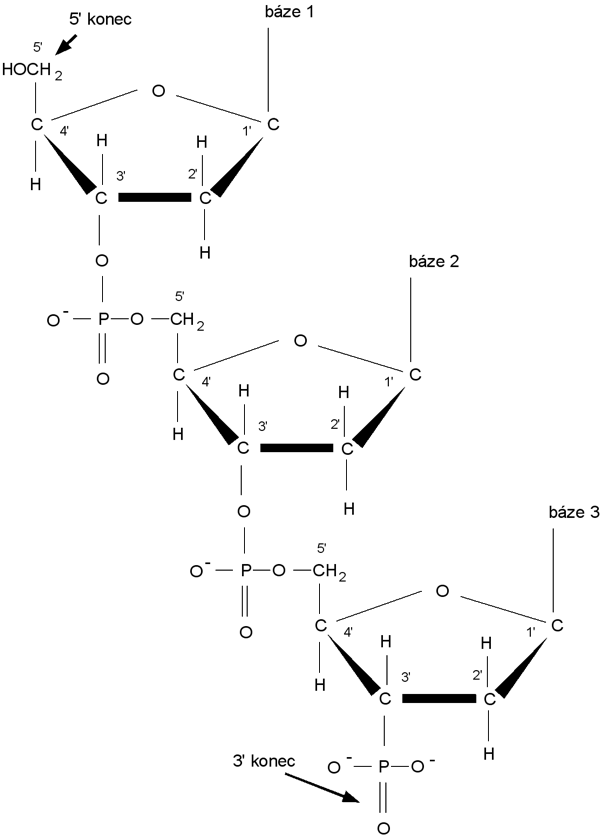
Část jednoho vlákna DNA tvořená třemi nukleotidy
Dvojice bází (A,T) a (C,G), se nazývají po objevitelích struktury DNA Watsonovy-Crickovy páry. Watsonovo-Crickovo párování významnou měrou přispívá k samoorganizaci nukleových kyselin, neboť díky elektronové komplementaritě struktury báze komplementárního páru fungují podobně jako zámek a klíč na molekulové úrovni. Toto párování je vysoce specifické, zajišťuje téměř bezchybný přepis sekvence bází z DNA do RNA, případně z rodičovské DNA do dceřiné DNA.
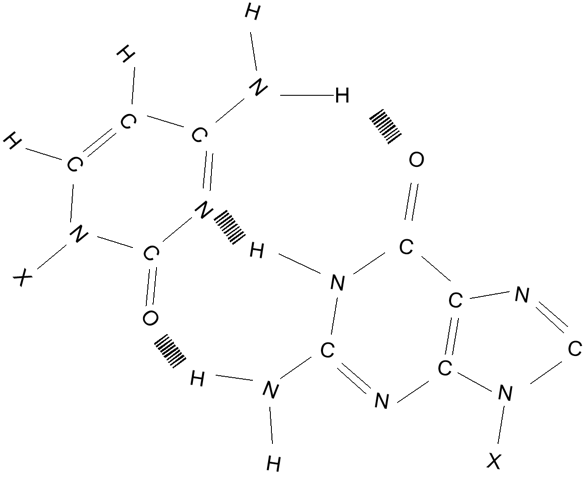
Párování komplementárních bází C-G, čárkovaně naznačeny vodíkové vazby.
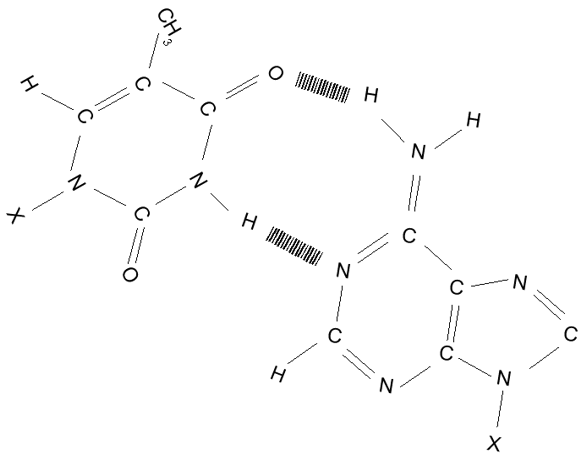
Párování komplementárních bází T-A, čárkovaně naznačeny vodíkové vazby.
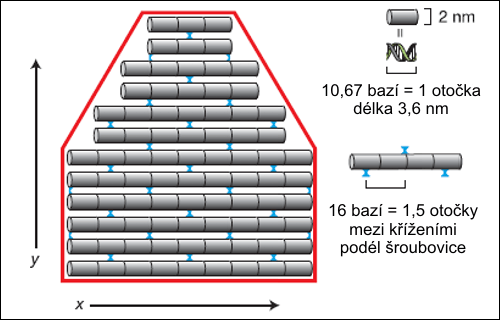
Ke každému vláknu DNA, označme si jej X, existuje jeho doplněk, komplementární vlákno, označme si jej Y. Ve vláknu Y báze navázané ve směru 3’–5’ odpovídají uspořádání komplementárních bází ve směru 5’–3’ vlákna X. Molekula DNA je tvořena dvojicí opačně orientovaných komplementárních vláken uspořádaných do dvojité šroubovice. Zevní kostrou každého vlákna jsou deoxyribózové cykly navzájem spojené zbytkem kyseliny fosforečné. Na cukerné složky obou koster navázané rovinné molekuly vždy jedné purinové a jedné pyrimidinové báze tvořící Watsonův-Crickův pár jsou uloženy prakticky kolmo k ose šroubovice. Dvojice bází se na sebe samovolně stohují podobně listům papíru ve středu vlákna, každá následující dvojice bází je pootočena o 36º. Průměr celého vlákna je zhruba 2 nm, na 3,4 nm stoupání závitu pravotočivé šroubovice DNA připadá deset párů bází. (Tato konformace DNA, B-DNA, není jediná možná, nicméně vytváří se za běžných podmínek v buňce. Při snížení relativního obsahu vody na 75 % B-DNA přechází vratně do konformačního uspořádání pravotočivé A-DNA, které je širší, má stoupání závitu 2,8 nm, 11 párů bází, a vytváří uvnitř DNA centrální dutinu. Roviny párů bází mají sklon 20º sklon vzhledem k ose šroubovice. Z-DNA je levotočivá se stoupáním 4,5 nm.)
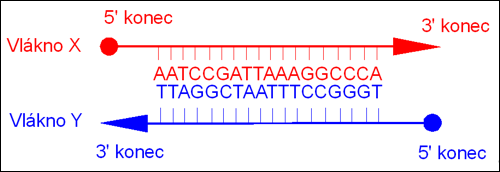
Komplementární vlákna dvouvláknové DNA.
Pro výrobu většího množství kopií DNA s požadovanou sekvencí nukleotidů je možné využít nitrobuněčných parazitů, virů nebo bakteriofágů. Je možné využít přímo jejich vlastní DNA, případně tato DNA může být pozměněna. Další možností namnožení většinou cizorodé DNA přímo v rychle se množících buňkách, obvykle bakteriálních. Jinou možností je namnožení DNA pomocí technických prostředků, například polymerázovou řetězovou reakcí PCR (Polymerase Chain Reaction). Jde o metodu zmnožení úseku DNA, ohraničeného dvěma známými sekvencemi bází. Reakční směs obsahuje primery, krátké, definované úseky jednořetězcové DNA, jejichž sekvence nukleotidů udává hranice zmnožovaného úseku, nukleotidy a polymerázu. Technika využívá rozpojení vláken dvouřetězcové DNA (dentaturaci, tání DNA) na dvě komplementární jednořetězcové DNA, zvýšením teploty. Po snížení teploty polymeráza naváže na primer a postupným doplňováním komplementárních nukleotidů dotvoří vlákna do dvouřetězových. Použitá polymeráza odolává zvýšení teploty, takže není nutné ji v každém cyklu do soustavy znovu dodávat. Jednotlivé kroky každého cyklu jsou řízeny teplotou reakční směsi. Počet úseků DNA exponenciálně narůstá s počtem cyklů. Typicky se používá zhruba 30 cyklů, které zmnoží požadovaný úsek miliardkrát.
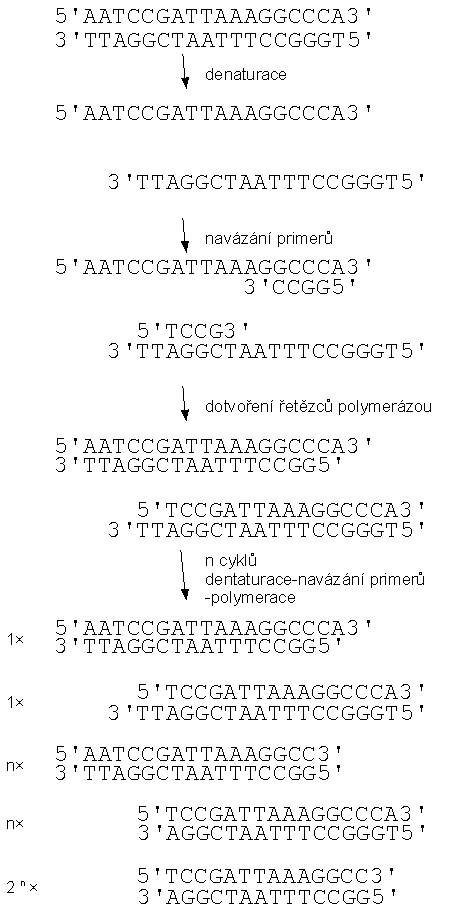
Schematický průběh PCR
1D, 2D, 3D…a je tu truhlička!
V buňkách se DNA přirozeně organizuje do lineárních struktur tvořených dlouhými tenkými vlákny. V zájmu uchování genetické informace je důležité uchování jednorozměrné struktury dvou po celé délce vzájemně komplementárních vláken. Co se však stane, pokud k jednomu dlouhému vláknu DNA přidáme kratší úseky DNA, které jsou částí komplementární jednomu úseku DNA a druhou částí jinému, vzdálenějšímu, navíc v opačném pořadí? Na dlouhém řetězci se vytvoří smyčka a kratší vytvoří příčný můstek mezi dvěma dvojšroubovicemi. Takto lze DNA skládat a vytvářet z ní obrazce podobně jako japonské skládačky origami z listu papíru.
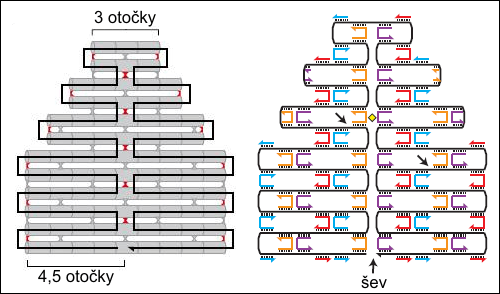
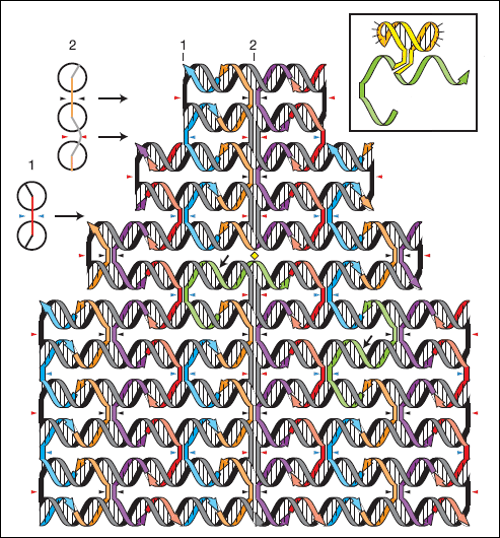
Vytváření dvojrozměrných struktur pomocí DNA. V schématech jsou
komplementární
úseky vyznačeny čárkovaně. Zdroj: Nature 440, 297-302 (16 March
2006).

DNA origami - dvourozměrné struktury vytvořené pomocí DNA.
Zdroj: Nature 440, 297-302 (16 March 2006).
Jakmile je možné z DNA vytvářet dvojrozměrné útvary, je to jen krůček ke spojení do třírozměrných objektů. Spojíme-li stěny krychle nebo kvádru v hranách, k čemuž opět můžeme použít kousky jednovláknové DNA, které je částí komplementární úsekům DNA na jedné stěně a druhou částí úsekům na druhé stěně, dostaneme dutou krabičku, které už do truhličky zbývá jediné – otevíratelné víko se zámkem. Jako zámek nám opět poslouží DNA. Dvě vlákna, každé ukotvené v jedné stěně, jsou navzájem komplementární. Jedno je však delší a vystavuje „lepivý konec“ příchozímu klíči, kterým, jak jinak, je DNA, jednovláknová a komplementární k delšímu vláknu zámku. Po vytlačení kratšího vlákna zámku klíčem lze otevření víka docílit vhodným pnutím „DNA pantů“ na hraně víka protilehlé k zámku.

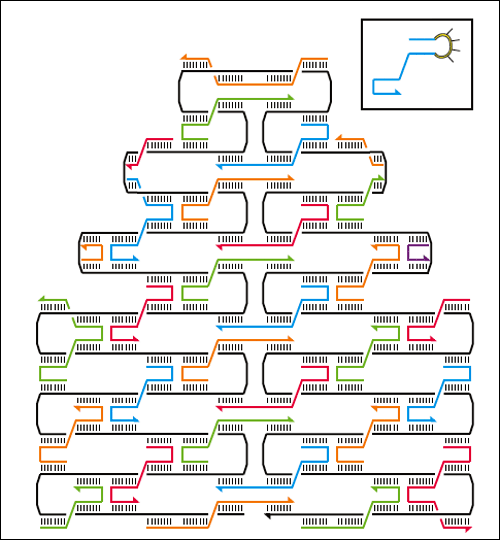
Rozložené stěny DNA truhličky vytvořené jako DNA origami. Šedé vlákno je tvořeno jedinou prstencovou jednořetězcovou DNA, v rozích stěn na sebe ve skutečnosti navazuje prostřednictvím spojek o 33 nukleotidech. Zdroj: Nature 459, 73-76 (7 May 2009).
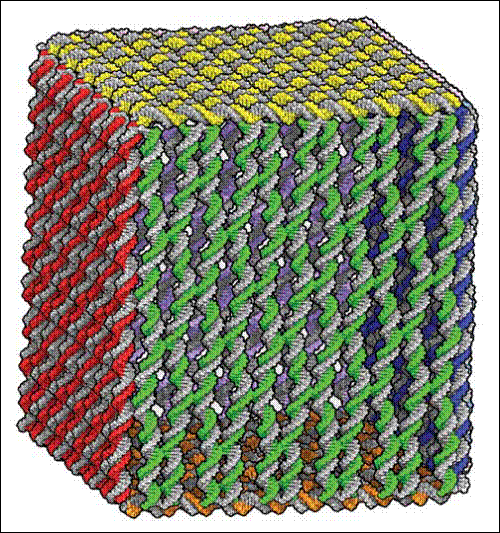
DNA origami složené do truhličky. Zdroj: Nature 459, 73-76 (7 May 2009).

AFM snímek DNA origami rozložených stěn truhličky.
Zdroj: Nature 459, 73-76 (7 May 2009).

AFM snímek DNA origami složené do truhličky.
Zdroj: Nature 459, 73-76 (7 May 2009).

AFM snímek DNA origami složené do truhličky s otevřeným víkem.
Zdroj: Nature 459, 73-76 (7 May 2009).
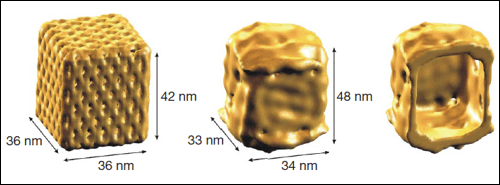
Teoretický model (vlevo), rekonstrukce povrchu z kryoelektronové
mikroskopie
(uprostřed) a řez dutinou (vpravo) nanotruhličky z DNA.
Zdroj: Nature 459, 73-76 (7 May 2009)

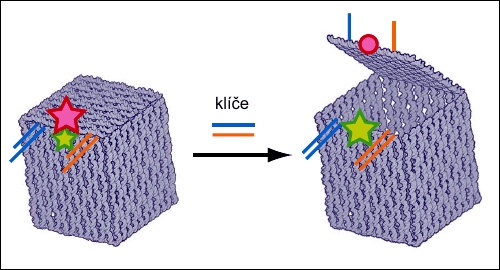
Kontrola otevření víka pomocí fluorescenčních barviv. Do DNA stěn se zabudují barviva, která při vzájemném přiblížení a oddálení mění svá spektra (uzavřená nanotruhlička svítí červeně, po otevření svítí zeleně podobně elektronickému zámku na dveřích či semaforu). Na obrázku je rovněž schematicky naznačen DNA zámek truhličky. Zdroj: Nature 459, 73-76 (7 May 2009).
Závěr
Inu, není nad to mít truhlu. Můžeme z ní udělat Pandořinu skříňku pro nádor, neboť jen jeho DNA otevře zámek a uvolní jedovatý obsah, můžeme si v ní dopravovat léky na místo určení a zmírnit či odstranit vedlejší příznaky, můžeme do ní zavírat a jinde vypouštět nežádoucí látky, ale můžeme se na ni dívat i jako na základ DNA počítače nové generace.
Animace týdne: Svinutí DNA do chromozómů během dělení buňky
Svinutí DNA do chromozómů během dělení buňky. Při pohledu na vlákno dvoušroubovice DNA jsou dobře patrné hluboké a mělké žlábky. Modře jsou znázorněny molekuly speciálních bílkovin, histonů, na které se DNA navinuje. Společně vytvářejí strukturu zvanou nukleozómy, které dalším překrucováním DNA vytvářejí chromatim. Svinování DNA do chromozómu lze přirovnat ke kroucení dlouhého gumového vlákna. Nejprve na vláknu vytvářejí smyčky, posléze vzniká tlustější vlákno tvořené smyčkami uloženými těsně vedle sebe, dalším kroucením se vytvářejí smyčky na vláknu ze smyček, které posléze vytvoří další, tlustější vlákno, tvořené těsně uloženými smyčkami vlákna tvořeného těsně uloženými smyčkami a proces se může opakovat na další, vyšší úrovni. Kroucení DNA zajišťují speciální enzymy – gyrázy, které ve vybraných místech rozpojí oba řetězce DNA, otočí je a posléze zase spojí. Tím se v DNA vytváří torzní napětí, které napomáhá sbalení vlákna původně tlustého dva nanometry do struktury viditelné v optickém mikroskopu, jak je ukázáno při přesunu chromozómů do obou dceřiných buněk.
V druhé části je předvedeno vytváření kopií DNA uvnitř buňky. Jelikož vlákno DNA může být vytvářeno pouze ve směru od 5‘ konce k 3‘ konci, dotváří se na jednom vlákně druhý řetězec průběžně, zatímco na druhém se musí tvořit po částech ve směru proti pohybu rozpojovaného vlákna. Zdroj: Drew Berry, The Walter and Elisa Hall Institute of Medical research, 2009. (Avi/xvid+mp3, 19 MB)
Odkazy
Andersen E.S. et al.: Self-assembly of a nanoscale DNA box with a controllable lid; Nature 459, 73-76 (7 May 2009)
![]()
![]()
